Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.5.7.2.4 Organelle Membrane Breach
Immotile submicron particles appear generally mechanocompatible with the intracellular environment. For example, the presence of artificial intracellular particles 20-200 nm in diameter (used as intracellular fluorescent labels or sensors) is not mechanically disruptive to the cell [4238, 4258]. Intracellular alumina particles can elicit changes in intracellular elemental composition and a reduction of phagocytic ability in human macrophages [2596], but there is no evidence that this is a mechanical effect. Silicon microdisks introduced into rabbit white cells were subsequently transported to a site of injury, with no evidence of functional cell impairment during cytocarriage [4259].
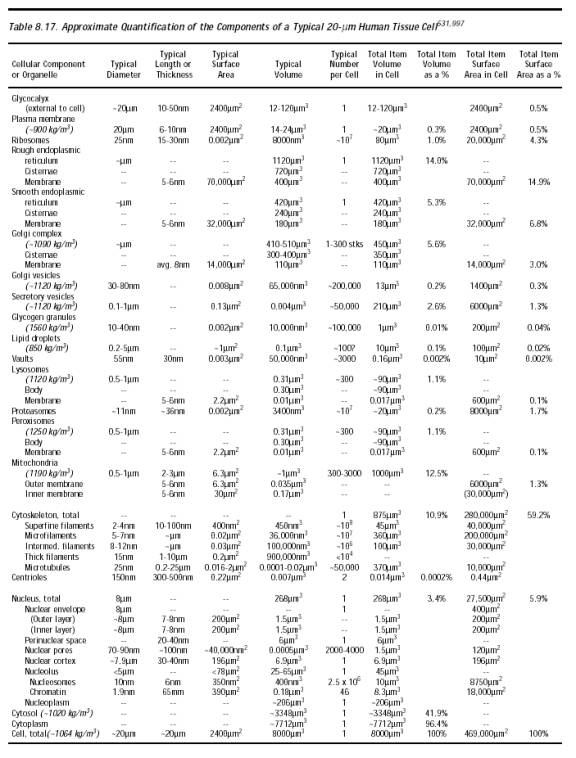
The plasma membrane (Section 8.5.3.2) represents only a tiny fraction of the total membrane present in the cell. Indeed, 99.5% of the cell’s lipid bilayer surface lies elsewhere (Table 8.17), mostly in intracellular organelles. The membranes of each type of organelle are structurally distinct (Section 8.5.3) and have different disease susceptibilities [4260]. But all should possess roughly comparable mechanical strength [4261-4263], so puncture forces should be similar as for plasma membrane. Because the Golgi complex is constructed via an ordered merger and coalescence of isolated intracellular vesicles, and readily reconstitutes itself from a vesiculated state [4264], it is unlikely that the interposition of a small passive nanorobot into these dynamic structures could mechanically influence their growth. A motile nanorobot locomoting around and through these structures could prove temporarily disruptive, though the structures would probably self-repair as suggested by the 10-minute recovery time for Golgi membranes that have been mechanically disrupted by the forcible interposition of artificial vesicles into the membranes under high g-force centrifugation [4265]. There is some evidence [4266] that the endoplasmic reticulum (ER) may be more sensitive to mechanical membrane damage [4266] or to the physical disruption of the supporting microtubule lattice [4267]. However, the integrity of the ER appears to be maintained during mitosis, with little or no fragmentation and vesiculation [4304].
The greatest threat to cell viability from organelle membrane rupture and intraorganelle leakage probably comes from the mitochondrion. The careless (or even purposeful) nanorobotic breaching of the integrity of this organelle [5672] could result in the cytosolic release of harmful mitochondrial proteins from the intermembrane space including especially cytochrome c, a 12.3 kD protein that forms part of a complex which directly activates caspase-9 [4268], one of the apical enzymes responsible for the apoptotic dismantling of the cell [4269]. A cytosolic concentration of ~0.1 µM (~0.01 pg/cell) of cytochrome c appears sufficient to trigger apoptosis [4270], but the human body contains ~3 mg of iron as cytochrome c [4271] distributed throughout the mitochondria of ~4 x 1012 tissue cells (Section 8.5.1), or ~0.20 pg/cell assuming ~20-micron cells. So at least ~5% of all cellular mitochondria probably must be fully ruptured by the mechanical activities of nanorobots, or an equivalent amount of intraorganelle fluid leakage must occur, in order to induce apoptosis, which seems an unlikely event. (Apoptosis may be intentionally aborted by in cyto nanorobots; Section 15.5.7.6.) Proteins attempting to enter the organelle that become stuck across both mitochondrial membranes “jam” protein import sites but do not collapse the potential across the mitochondrial inner membrane [4272].
What about lysosomes? Although it is normally quite stable, the lysosome membrane can become more fragile: (1) when the cell is injured [4273] or deprived of oxygen [4274]; (2) when excessive amounts of vitamin A (hypervitaminosis A [4275]) or iron [4276] are present; (3) during long-term exposure to gentamicin (though with no increase in cell mortality [4277]); or even (4) in some cases of congenital vascular anomalies [4278]. Lysosomes may undergo drastic shape changes during microautophagocytosis, including invagination of their boundary membrane with scission of vesicles into the lumen of the organelle [5482]. Tubular lysosomes 75 nm in diameter and 2-3 microns long extending outward from the nucleus are common in macrophages, and are fragmented if the supporting microtubule lattice is physically disrupted [4279]. However, if left alone without further disturbance, the fragmented microtubules reassemble and the tubular lysosomes reappear within 10-20 minutes [4279].
What about mechanical stress damage? One experiment [4280] found possible lysosomal membrane damage in myocardial swine cells from animals subjected to high +Gz accelerations. But another experiment [4281] found that mechanical traction strain applied to cultured retinal pigment epithelium cells and fibroblasts sufficient to break intercellular attachments does not disrupt lysosomal membrane integrity during stretching. Post-phagocytosis Listeria monocytogenes bacteria exit the lysosome in which they reside by penetrating through the lysosomal wall, a mechanical disturbance that does not immediately impair host cell function [4282].
Lysosomes were once called “suicide sacs” because lysosomal rupture can result in self-digestion of the cell, a process known as autolysis. But it is now known that lysosomes are part of the normal cellular digestion apparatus relating the process of endocytosis to the processes of intracellular synthesis, storage, and transport [1767], even including intraorganelle vesicles [4283]. Structural deterioration of lysosomes does not occur rapidly in ischemic or postmortem cells [4284, 4285] or even in cells subjected to microwave irradiation [4286]. Direct damage from organelle fluid leakage, should such leakage occur, may be minimal because most of the digestive enzymes in lysosomes require the relatively low pH of the lysosomal vesicles for activation – just as some proteases require the low pH of the stomach. Peroxisomal membranes appear equally durable [4287]. Lysosomal membrane rupture could release limited amounts of lipofuscin into the cytosol since these organelles are the primary site of lipofuscinosis [4288], which would probably be reasonably harmless (Section 15.6.3.2).
What about the nuclear membrane? Nuclear-cytoskeletal [4289] manipulations that alter the cellular mechanical force balance can cause the nucleus to change shape [4290, 5445] and nuclear envelope fragility increases in the presence of high concentrations of salt [4291]. But micropipette injection of DNA material into cell nuclei is a common biotechnology procedure [4292-4295] which is easily survived by the cell. Normally, the nuclear envelope is reversibly disassembled and reassembled during mitosis, a highly regulated process [4296, 4297] that includes the physical tearing apart of the nuclear envelope by extranuclear microtubules that penetrate it [4304]. Reassembly of a nuclear envelope that has been mechanically disrupted, fragmented, or completely disassembled by arbitrary artificial external forces (e.g., due to perinuclear nanorobotic activities) outside of the normal mitotic cycle cannot rely upon the regulated sequential cell cycle process [4298-4304] to guide reassembly. Such disruptions possibly might lead to apoptosis (Section 10.4.1.1), especially if the native chromosomes or their intranuclear moorings are physically damaged in any way [4421, 5465-5467], or to chromatin digestion – e.g., both single- and double-stranded circular plasmid DNA has a 50-90 minute half-life in mammalian cytosol probably due to cytosolic nucleases [4295, 4305, 4306]. In some circumstances it may be possible for in cyto medical nanorobots to induce reassembly of the disturbed nuclear membranes [4307]. Basic restrictions on the speed of mechanical motions that may safely be applied to chromatin are briefly discussed in Section 8.5.4.7.
Last updated on 30 April 2004
{kind=link}