Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.5.7.3.2 Disruption of Molecular Motors and Vesicular Transport
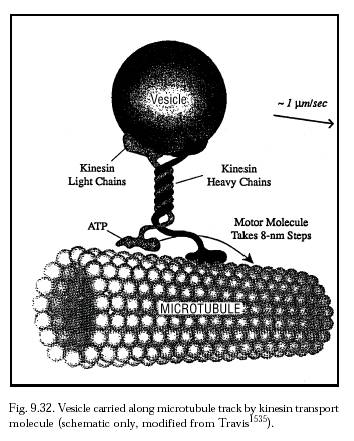
There are a variety of human disorders associated with dysfunction of cytoskeleton-based molecular motors [4351], including, for example, the motor-based diseases involving defective cellular myosin motors [4352-4354] (e.g., implicated in Griscelli syndrome [6219], hearing loss [6220], hypertrophic cardiomyopathy [4352], and other myosin myopathies [6221]), spindle assembly- and function-related diseases [4355] or kinesin- and dynein-related motor diseases (e.g., implicated [6222] in Charcot-Marie-Tooth disease type 2A [6223], Kartagener syndrome [6224] or primary ciliary dyskinesia [6225], lissencephaly [6226], polycystic kidney disease [6227], and retinitis pigmentosa [6228, 6229]), and other avenues for cellular malfunction [4356, 4357, 6222, 6230]. Cellular motors also participate in the self-organization of microtubule network structures [4347]. But perhaps the most important function of molecular motors is intracellular vesicular transport, and most particularly axonal transport in neurons [4358, 4359, 6231-6234]. Typically, organelles, vesicles and granules ~100 nm in diameter or larger are carried at a peak speed of up to ~2 microns/sec on the back of a 60-nm kinesin transport molecule (Figure 9.32) that takes 8-nm ATP-powered steps along microtubule tracks running throughout the cell [4360-4364]. (Mean unloaded kinesin motor speed is usually only 0.5-0.8 microns/sec.) Conventional kinesin is a dimer of identical ~120 kD protein chains [4371] with a diffusion coefficient D ~ 2 x 10-11 m2/sec [4347]. The vesicle-attached kinesin motor molecule steps toward the plus-end of microtubules by converting the energy of ATP hydrolysis to mechanical work.
Could a vesicle be dislodged from its microtubule track after being bumped by a passing intracellular nanorobot? A kinesin molecular mechanical detachment force of Fdetach ~ 13 pN [4366] requires that a 1-micron3 diamondoid nanorobot of mass mnano ~ 2 x 10-15 kg must impact and carry the cargo vesicle a distance Svesicle ~ 1 nm to detach it at a constant velocity of vnano >~ (2 Svesicle Fdetach / mnano)1/2 ~ 3500 micron/sec, well above the self-imposed 10 micron/sec intracellular locomotion speed limit (Section 15.5.7.3.1) and even slightly exceeding the nanorobot instantaneous thermal velocity of ~2500 microns/sec in water at 37 oC (Eqn. 3.3). The torsional stiffness of kinesin is so low that the molecule readily twists through more than 360o from its resting orientation [4368], thus allowing the cargo to easily swivel out of the way of foreign impacting objects. And kinesin motors normally detach from a microtubule after a few seconds of travel anyway [4369-4371]. Still, nanorobots should be able to exert mechanical forces well in excess of 13 pN, so care should be taken to minimize those motions and trajectories which are likely to produce kinesin detachment. Analogously, it has been found experimentally that intracellular microspheres experience enhanced diffusion over short time scales near the nucleus, possibly due to interactions with microtubule-associated motor proteins [4373].
Could a nanorobot that has clamped or securely bound itself to one location on a microtubule prevent the passage of vesicular cargoes, causing vesicles to bunch up behind or to detach? And could kinesin motor molecules that encounter the obstruction be permanently damaged? Most likely a detached vesicle will reattach to a clear neighboring microtubule and continue its trek [4374, 4375], or will reattach to the original microtubule downstream of the nanorobotic obstruction. In some cases organelles can attach to and move along multiple filaments simultaneously [4376] using multiple motor molecules [4377], potentially reducing the interference with forward motion from a single-filament blockage.
Coppin, Pierce, Hsu and Vale [4366] have carefully studied the behavior of kinesin molecules whose forward progress is mechanically constrained. Kinesin has a stall load of ~5 pN [4364-4367]. There is an increasing rate of dissociation with increasing load. Specifically, the dissociation rate is ~0.2/sec at 1 pN load, 0.5/sec at 2 pN, 1/sec at 3 pN, and 2/sec at 5 pN load [4366], rates which can be altered by the presence of microtubule-associated proteins [4378]. However, super stall loads of 5-13 pN do not cause kinesin to walk backwards, “probably because of an irreversible transition in the mechanical cycle.” Rather, when super-stalled the kinesin motor most commonly takes a single backward movement and then dissociates (detaches) from the microtubule,* occasionally rebinding to the same microtubule (always at/below the stall load) and resuming its normal movement. This clearly demonstrates that the kinesin motor is still functional after being subjected to a dissociative induced stall. That is, a superload-induced detachment doesn’t “break” the motor. The conclusion is that similar mechanical interference by a nanorobot also should not damage a processive protein motor. Interestingly, a mutant form of kinesin with its ATP and microtubule binding sites decoupled has been found that binds so tightly to the microtubule that the motor cannot let go [4379], crudely analogous to the case of a nanorobot that firmly grasps a microtubule for a period long in comparison to the timescale of kinesin procession.
* Alternating back and forth movements also are observed at super-stall [4366]. The dissociation rate increases with load as long as the motor is moving (up to 2 Hz at 5 pN), but then becomes independent of load once the motor stalls – e.g., the stall time is 0.57 sec, representing a dissociation rate of ~1.8 Hz, for either spontaneous (~5 pN) or induced (~12 pN) stalls. Interestingly, forward loads induce the kinesin motor molecule to step faster under a wide range of ATP concentrations [4366]. Forward loads of 5 pN increase velocity by +200% if ATP concentration is rate-limiting (5-40 µM) or by +50% if ATP is saturated (1 mM), but forward loads >5 pN cause forward velocity to drop off sharply [4366]. A small carboxyl domain acts as a switch that turns the motor off when the kinesin motor is not bound to cargo [4380].
A nanorobot ambulating along microtubules should endeavor to avoid applying lateral forces exceeding the kinesin detachment load of ~13 pN [4366] which could have the effect of detaching associated vesicles as the nanorobot progresses, like a tree limb being stripped of its leaves as it is pulled through a tight-fitting metal ring. (Using a two-dimensional optical force clamp, researchers in Block’s group [6172] observed that ambulating kinesin molecules subjected to sideways forces up to 8 pN only slowed by ~30%, and similar forces applied from the rear have only a weak effect on forward speed.) Typically, the processive kinesin molecule only takes a few hundred steps before letting go [4369-4371], so an occasional early detachment, by itself, should not induce cellular dysfunction. As long as the processive motor protein is not physically damaged, most of the detached vesicles should reattach and continue their journey after the nanorobot has passed by. The minimum spacing (maximum density) of kinesin motors along a microtubule is an axial repeat of 8 nm [4381] and experiments with isolated microtubules gliding on kinesin-coated glass find optimal motility at a 47 nm separation between kinesins [4382]. But the vesicular transport of 100-2000 nm diameter organelles in cyto implies that propulsive kinesin contacts are normally spaced at least 0.1-2 microns apart along the microtubule tracks. This leaves plenty of unoccupied foothold space to allow nanorobots to avoid disrupting processive motor proteins already in transit.
Coordinated groups of in cyto medical nanorobots should avoid inadvertently corralling or bulldozing large numbers of vesicles or motor molecules into relatively small volumes within the cell, as such increases in motor molecule concentration could increase the local microtubule polymerization rate [4347]. Analogously, in motor neuron diseases where vesicular transport is blocked by massive localized accumulations of kinesin molecules, the blockage can produce large axonal swellings in the motor neurons in human spinal cords and can disturb the machinery for anterograde fast axonal transport [4383]. Intracellular traffic jams involving repositioned vesicles and organelles [4384] appear to be initiated by the accumulation of stalled kinesin cargoes [4385] and are most commonly reported in neurons [4386-4389] where their effects are most serious. For example, axonal organelles transported by kinesin molecules that stall can cause organelle jams that disrupt retrograde as well as anterograde fast axonal transport, leading to defective action potentials, dystrophic terminals, reduced transmitter secretion and progressive distal paralysis that parallels the pathologies of motor diseases such as amyotrophic lateral sclerosis [4388]. Stretch injury to axonal cytoskeleton resulting in major loss of microtubules disrupts fast axonal transport resulting in focal accumulation of membranous organelles and axonal swellings [4387], and a chemically-created microtubule-free region can serve as a trap that causes axonally transported particles to accumulate into a swollen region [4386]. Nanorobots should avoid creating such regions within the cell.
Similar considerations also apply during operations by intranuclear nanorobots, given the presence of myosin-based motors [4390], RNA polymerase motors [4391], and other motor molecules inside the cell nucleus.
Last updated on 30 April 2004
{kind=link}