Nanomedicine, Volume I: Basic Capabilities
© 1999 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume I: Basic Capabilities, Landes Bioscience, Georgetown, TX, 1999
4.2.1 Broadband Receptor Arrays
One simple broadband concentration sensor, shown schematically10 in Figure 4.1, consists of a graduated series of receptors having uniformly high specificity but engineered with progressively greater affinities (successively smaller equilibrium dissociation constants Kd (molecules/nm3); Section 3.5.2) for the target molecule, which is present in the test sample at concentration cligand. Exposure of the receptor array to the test sample for a time tEQ necessary to reach diffusion-driven equilibrium gives a probability of receptor occupancy Pocc ~ 0.91 in a receptor with Kd ~ 0.1cligand, Pocc ~ 0.50 in a receptor with Kd ~ cligand, and Pocc ~ 0.09 in a receptor with Kd ~ 10cligand.
During each measurement cycle, all steric probes are simultaneously extended into their associated receptor volumes at a time tEQ after presentation of sample to the array. Probes which reach full extension register an empty receptor; those which cannot fully extend register an occupied receptor. After registration, the probes are retracted and ejection rods are thrust into all receptors (typically requiring ~1 nanonewton (nN) of force for occupied receptors) to empty them while the test chamber is flushed clear in preparation for the next cycle to begin. Ideally, the probe rod pushing into the binding site should push the bound molecule further into the binding site to avoid complications that may arise due to competing kinetic barriers, and ejection rods should push molecules far enough to ensure that the next resample is independent.
Consider a series of N steric probe units as proposed by Drexler.10 Each probe measures 8 nm x (2.5 nm)2 ~ 50 nm3 and has mass ~ 2 x 10-22 kg. The ratio of the dissociation constants of adjacent units is k = Kdi / Kdi+1 > 1, with Kd1 ~ 10-4 molecules/nm3 and KdN ~10-13 molecules/nm3 typically in the human body, and so N = 1 +{log10 (Kd1/KdN) / log10 (k)}.
Minimum measurement error occurs when cligand exactly matches the Kd of a probe unit, that is, when cligand = Kdi (i.e., Poccupiedi = 0.5). Maximum measurement error occurs when cligand lies exactly midway between two probes such that Kdi > cligand > Kdi+1, or, more specifically, at the geometric midpoint cligand = (Kdi Kdi+1)1/2 = Kdi k-1/2. In this case, Eqn. 3.24 becomes
where Punoccupiedi = 1 - Poccupiedi. Additionally, sampling error is ~Nm-1/2 when Nm independent measurements are taken (i.e., Nm measurement cycles employing all N probe units during each cycle), for Nm >> 1, establishing an error bound on the probability of receptor occupancy of Poccupiedi ± Nm-1/2. If two concentrations c1 = Poccupiedi / (1 - Poccupiedi) and c2 = (Poccupiedi + Nm-1/2) / (1 - (Poccupiedi + Nm-1/2)) may be distinguished, where Poccupiedi is given by Eqn. 4.1, then the minimum detectable concentration differential Dc /c is
For k >> 1, Eqn. 4.2 reduces to: (Dc / c)large ~ (1 + k1/2) / Nm1/2. In the limit as k approaches 1 and Nm >> 1, Eqn. 4.2 reduces to: (Dc / c)small ~ k-1/2 (Dc / c)large.
For k = 10 (an N = 10-probe sensor, sufficient to span the entire Kd = 10-4 to 10-13 molecules/nm3 range using one probe per decade), Dc / c = 0.94 (94%) at Nm = 100, 0.20 (20%) at Nm = 1000, (0.06) 6% at Nm = 10,000, and 0.006 (0.6%) at Nm = 106 measurement cycles. Little additional discrimination is obtained by using more than N ~ 10 probes in the sensor. For Nm = 1000, Dc / c = 0.89 (89%) at k = 178 (N = 5 probes), 0.20 (20%) at k = 10 (N = 10 probes), 0.136 (13.6%) at k = 1.23 (N = 100 probes), and 0.135 (13.5%) at k = 1.0021 (N = 10,000 probes). Note that Dc / c = 0.20 (20%) is equivalent to detecting a pH variation of ~0.08 (e.g., 10(0.08) - 1 ~ 0.20).
Sensor cycle time ~tEQ may be crudely approximated by the number of random ligand-receptor encounters (Nencounters ~ 100; Drexler10) necessary to ensure binding divided by the number of ligands striking the receptor surface per second,434 or (sec/cycle)
where A = active receptor cross-sectional area ~0.1 nm2 for small molecule (MW ~100 gm/mole) receptors to ~10 nm2 for large molecule (MW ~105 gm/mole) receptors; cligand = 10-2 nm-3 for the most common molecules to 10-12 nm-3 for rare molecules in the human body; MWkg = target ligand molecular weight in kg/mole, k = 0.01381 zJ/K (Boltzmann constant), T = 310 K (human body temperature), and NA = 6.023 x 1023 molecules/mole (Avogadro's number).
For common small molecules, tEQ ~10-6 sec; for common large molecules, tEQ ~0.2 x 10-6 sec, but a sample chamber ~(25 nm)3 in size is diffusion-limited to a minimum tEQ ~ 10-5 sec for cligand >~ 10-4 nm-3. For the rarest small molecules in the human body, tEQ ~ 800 sec, ~200 sec for rare large molecules, and a larger chamber may also be required. Preconcentration of the test sample by dehydration using sorting rotors (Section 3.4.2) to rapidly remove water from the sample chamber reduces tEQ to 10-40 sec for very rare molecules.
Steric probes using a driving force10 of ~100 pN, over 2 nm of motion per cycle require 200 zJ/measurement. If 1000 measurement cycles of a 10-probe sensor suffice to determine concentration to ~2 significant figures, this information costs ~0.002 pJ to acquire, a power consumption of from ~10 pW for large common molecules to ~10-9 pW (~0.6 kT/sec) for small rare molecules while the sensor is working. Power dissipation may run up to ~10 times higher, depending upon the number of occupied receptors that must be cleared via ejection rods during each cycle.
Last updated on 16 February 2003
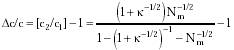 {Eqn. 4.2}
{Eqn. 4.2}