Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.3.3.5 Amorphous Carbon Particles
It has already been noted that finely divided amorphous carbon particles generally are well tolerated by the body [181, 632, 850]. The essentially passive nature of carbon in human flesh has been known since ancient times, and India ink [778], charcoal and lampblack (roughly spherical 10-20 nm particles) have been used for ornamental and official tattoos [516]. Colloidal carbon is usually assumed to be nontoxic [773, 4873]. Large amounts of carbon particulate debris loose in the body, even around the spinal dura and nerves [839], evidently are tolerable. Carbon black (CB), the most common amorphous carbon particles that have been subjected to extensive biological experimentation, are regarded as largely inert [851]. Carbon black is distinctly different from, and more benign than, carbon soot [4867]. Ultrafine carbon black particles are typically ~14 nm in diameter, while ordinary “fine” carbon black particles are typically ~260 nm in diameter [769]. Commercial carbon blacks may contain 88-99.5% carbon, 0.3-11% oxygen, 0.1-1% hydrogen, up to 1% inorganic material, small amounts of other organics, and traces of sulfur, depending upon the method of manufacture. Both CB and ultrafine CB may contain numerous biologically relevant metals. One assay [769] of ultrafine CB found 19 ppm Fe, 11 ppm Zn, 3.5 ppm Cr, 1.9 ppm Pb, 0.8 ppm Cu, 0.02 ppm Be, and even 0.0039 ppm Tl, all of which may enhance the biological activity of these particles.
India ink, traditionally used for drawing, is a dispersion of carbon black in water. (One MSDS for India ink [6081] lists its only health hazard as irritant with TLV TWA of 2 mg/mm3.) The suspension is often stabilized by various alkaline solutions, shellac dissolved in borax solution, soap, gelatin, glue, gum arabic, or dextrin. India ink is often used as a phagocytosis labeling agent or a cell differentiation tracer because CB is easily administered, is relatively nontoxic, and is easily observed in cells that have taken it up. India ink is useful diagnostically for detecting cryptococcal meningitis [5918], easily demonstrating individual cryptococcus organisms by phase contrast microscopy. CB is also employed in tattoos, both cosmetic and medical. For example, India ink is used for endoscopic colonic tattooing [852-856, 4861] and lymphatic mapping [4862]. It produces a long-lasting stain “with relatively low risk of adverse reaction and toxicity” [856]. There are just a few rare cases reported of allergic reaction [855] to India ink, fever and abdominal pain [856], colonic abscess and focal peritonitis [853], abscess-like pseudotumor with chronic granulomatous inflammation [856], inflammatory infiltrate with microhemorrhage and thrombosis [852], inflamed vessels but without fibrinoid necrosis [854], and early reactions including necrosis, edema, and neutrophilic infiltration in the submucosa and muscularis propria [854]. M. Sprintz notes it is possible that these adverse reactions were caused by something other than India ink, such as microperforations in the bowel during the procedures.
The chemical character of the CB particle surface mediates its biological reactivity [857]. The often partially-saturated attractive forces can allow these surfaces to readily adsorb large amounts of gases and solutes from solution [858]. Active surface groups on carbon blacks have been shown to adsorb and retain gaseous adsorbate molecules selectively [859, 860]. Histone adsorption on the surface of carbon particles significantly stimulates their ingestion by rat peritoneal macrophages, hamster kidney fibroblasts, and mouse L-cells [861], and the adsorption of polycyclic aromatic hydrocarbons (PAHs) and their nonpolar metabolites on the surface of carbon particles ingested by rat alveolar macrophages has been studied [862].
The carbon immunoassay (CIA) [863] is a direct serological test relying on a specific reaction between the carbon particles of India ink and rabbit immunoglobulin G. (This assay is also known as the India-ink immuno-reaction (IIR) test [864-866].) The carbon particles must be pretreated with staphylococcal protein A to induce a reaction to IgG antibodies in human serum in tests for toxoplasmosis. Carbon black can also have a significant adjuvant effect on the local immune-mediated inflammatory response and on the systemic specific IgE response to allergen (ovalbumin) [867]. When CB is administered along with the allergen to mice, there is a significantly augmented response in the draining popliteal lymph node including increases in weight, cell numbers, cell proliferation, and local lymph node response duration [867]. Carbon black immunochromatographic testing is well known [5882].
Immune and other cells clearly respond to the presence of carbon particles placed in the body. In one in vivo study [868], chicken basophils, neutrophils, monocytes and platelets showed phagocytosis of dermally-implanted carbon particles although eosinophils ingested these particles only occasionally. In another chicken study [883], platelets and monocytes took up carbon particles but neutrophils did not. In a human study [869], scattered wear particles from carbon-coated subperiostal implants were surrounded by lymphocytes, macrophages, plasma cells, and tissue eosinophils. Active phagocytosis and Russell bodies were seen, with large masses of carbon surrounded by connective tissue [869]. In another study using cultured human monocytic cells [4857], carbon black particles or diesel exhaust particles, by themselves, were unable to induce HLA-DR when applied to the surface of THP-1 antigen-presenting cells at concentrations of 0.1-1000 ng/cm2 after 48 hours of incubation. However, carbon black (>1 ng/cm2) plus diesel exhaust particles (>0.1 ng/cm2) interacted with IFN-gamma (a known HLA-DR enhancer) to increase HLA-DR expression up to 2.5-fold, indicating a surface-chemistry-related “adjuvant” effect. The possibility that nanorobots could accidentally serve as adjuvants to induce cell damage [4857] or immune system activity [4860, 4871] by other particles already present in the environment should be investigated further.
The uptake and long-term storage of carbon ink particles by the dermal and subcutaneous fibroblasts [778] is believed to represent a specific non-inflammatory defense mechanism that protects the living body, without immune reactions, against injuries and invasions by non-toxic foreign agencies. In young rats, subcutaneous injection of 0.02 ml of India ink almost completely eliminated small recirculating lymphocytes from the affected nodal structures, except in the center of the deep cortex units, similar to the effects of a whole-body 500 rad irradiation [870] (Section 6.3.7.1). Colloidal carbon particles injected into the cerebral cortex of the neonatal rat were readily ingested by young astrocytes [773]. Colloidal carbon administered intravenously in rats can retard chemotactic migration and phagocytic activity [871] or even produce complete blockade of the RES [872]. Macrophage blockade has been induced in mice by intravenous injection of ~1000 mg/kg of carbon particles [873]. Examination of the endothelial linings of capillaries, postcapillary venules and terminal arterioles also reveals a pronounced uptake of carbon particles by endothelial cells, different degrees of endothelial cell swelling, and often bulging into the microvessel lumens, possibly altering microvascular tone and arteriolar reactivity [872].
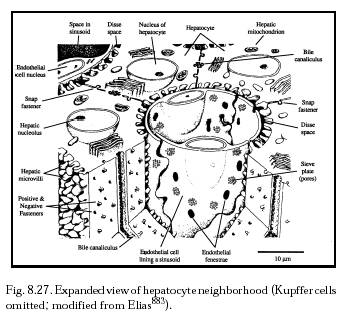
Circulating monocytes ingest intravenously-injected colloidal carbon particles [874, 883]. And uptake of intravenous India ink particles may transform monocytes into Kupffer cells [779]. In one experiment [777], Kupffer cells as well as hepatic sinusoidal endothelial cells took up Indian ink particles by pinocytosis, and a few Ito cells and hepatocytes ingested a small number of particles. One month after the injection, large clumps of aggregated Kupffer cells containing numerous carbon-filled vacuoles were distributed in the Disse space (Figure 8.27) and other connective tissue spaces. The Kupffer cells in these clumps were in close contact and were partly fused with one another. After 3-6 months, large multinucleate foreign body giant cells with numerous large vacuoles containing densely-packed ink particles were seen throughout the liver tissue, probably formed by the fusion of Kupffer cells [777]. Kupffer cells in aging mice show reduced phagocytosis of colloidal carbon [4863].
Macrophages are known to clear carbon particles from the blood [775]. Mouse omental macrophages phagocytize intraperitoneally injected carbon particles [876], and colloidal carbon particles injected intravitreously into chicken eyes are actively ingested by hyalocytes (the resident macrophages) by the second day, without significant leukocyte recruitment [771]. After 30 days the carbon-laden macrophages disappeared from the vitreous body but accumulated on the pecten oculi and retina [771], probably producing some small diminution of visual sensation. The number of free macrophages in mouse lungs increases in response to the intratracheal instillation of carbon particles [766]. There is early mononuclear phagocyte cell migration into alveoli and bronchioles from the blood compartment (e.g., from small pulmonary vessels [772]) and later migrations due to egression of interstitial lung macrophages [877] which exhibit increased mitotic activity [772]. This is observed for up to 6 months post-instillation [766], with macrophage accumulation localized to the areas of particulate deposition [767]. Ingestion of large amounts of carbon particles by alveolar macrophages also decreases the release of superoxide and hydrogen peroxide radicals [879] and reduces acid phosphatase and lysozyme enzyme intracellular concentrations [880] during bacterial phagocytosis. Elsewhere in the body, ingested insoluble carbon particles that accumulate in mouse intestinal Peyer’s patches do not produce detectable alterations in macrophage morphology [780], though carbon particles have induced production of macrophage colony-stimulating factor in mice [878].
Platelets are also known to clear colloidal carbon particles from the blood [775, 881-883]. For example, a single intravenous injection of colloidal carbon particles in Lewis rats at a dose of 320 mg/kg produced a prompt thrombocytopenia (reduced platelet count), with temporary sequestration of platelets in lung, liver and spleen [875]. Rat platelets were found to be involved in the initial removal of carbon from the blood [875]. Localization to the mesangium (the support structure of the renal glomerulus) is dose-dependent [875] and peaks at 32 hours post-injection [774]. India ink injected intravenously into mice also induces thrombocytopenia and markedly prolonged bleeding times (and prolonged thromboplastin and prothrombin times), reduces fibrinogen concentrations, and produces some cerebral thromboemboli, but does not cause substantial mast cell degranulation [884]. Chicken platelets also showed marked phagocytosis of carbon particles in both in vivo and in vitro systems [883]. Initially in both systems, platelets containing carbon particles tended to form clumps, but in the later stages clumping was less obvious in the in vivo system [883]. Another experiment described a simple and reproducible test for the phagocytic ability of human platelets [885]. Platelets obtained from heparinized venous blood were incubated with 0.11-micron colloidal carbon particles in autologous plasma at 37 oC. The number of platelets with or without carbon particles on smear preparations was determined at fixed intervals. Electron microscope observation confirmed that carbon particles were internalized in the vesicular or canalicular structures of about 67% of all platelets [885].
The responses of various organs to carbon particles have also been investigated experimentally. The eye seems particularly sensitive. In one experiment [886], 20-nm carbon particles were injected into the vitreous humor of rabbit eyes. Histological examination 8-10 weeks later showed partial posterior vitreous detachments, epiretinal cellular proliferation, and membranes in all eyes and retinal detachments in five eyes. Electron microscopy disclosed that the epiretinal membranes (resembling idiopathic preretinal gliosis or macular pucker) were formed mainly by Muller cell expansions, astrocytes, and macrophages. Muller cells penetrated the internal limiting membrane and removed carbon particles from the vitreous by endocytosis [886]. The experiments found that gaps are produced in the internal limiting membrane by glial cells and by macrophages that invade the vitreous in an attempt to remove foreign material.
In a subsequent experiment at the same laboratory [887], 20-70 nm carbon microparticles injected into the vitreous humor of cynomolgus monkeys induced intravitreal cellular proliferation. At 1 week, there was conspicuous cyclitis showing exudative separation of the nonpigmented and pigmented ciliary epithelium, inflammatory cells, mononuclear phagocytes, and premacular vitreous detachment. Continued macrophagic response was accompanied by fibrovascular proliferation with ingrowth of vessels from the ciliary body into the vitreous at 3 weeks. By 4-5 weeks there was deposition of extracellular fibrous material and traction retinal detachment. At 10 weeks, all eyes had extensive retinal detachment with pre- and subretinal collagenous cellular membranes. Carbon-laden macrophages were aggregated over the optic disc and fovea, and prepapillary neovascularization and cystoid macular edema was seen. The exposure of the interior of the eye to carbon particles thus produced an inflammatory and phagocytic response, which induced intravitreal fibrovascular proliferation, vitreous contraction, and retinal detachment [887] – a possibly cautionary result for long-term ocular-indwelling medical nanorobots of similar size.
The response of lung tissue to carbon particles has been extensively studied. For example, chronic inhalation of carbon black particles in air can produce carcinoma in rat lungs. Chronic [760, 4865] and subchronic [761] exposures impaired lung clearance and significantly increased mutation frequency in the hprt gene of rat alveolar epithelial cells at 7.1 mg/m3 and above, but produced no detectable adverse lung effects at a 1.1 mg/m3 level [760]. The official industrial threshold limit value for pure carbon black is 3.5 mg/m3 [764]. Inhalation of carbon black can produce pulmonary neoplasms in chronically exposed rats [888]. This is believed to be a result of a high lung burden of carbonaceous particles rather than from the genotoxicity of organic constituents [888]. Macrophages and neutrophils elicited by carbon black particles can exert a mutagenic effect on in vitro epithelial cells [889]. Carbon black has been linked to lung and bladder cancers at high occupational exposures [4864] (though not confirmed [4870]) and possibly also to genotoxicity in human alveolar epithelial cells [4869] and in rat lung tumors [5881]. Ultrafine carbon black particles activate apoptosis-related pathways in alveolar epithelial cells, whereas fine carbon black does not [4858]. At least one study claims that inhaling carbon black may transiently elevate the risk of myocardial infarctions in humans within a few hours after exposure [4866].
Inflammatory effects of carbon black particles on the lungs are well studied [890]. For instance, rats intratracheally instilled with saline suspensions of 10 mg/kg or 100 mg/kg doses of carbon black produced neutrophilic inflammation in all rats at both doses [889]. Epithelial hyperplasia and elevated hprt gene mutation frequency in alveolar Type II cells were observed only at the higher dose [889]. Carbon particles instilled into the lungs of mice induced an inflammatory response with excess production of alveolar macrophages for 2 weeks, after which the macrophage count returned to normal with normal lung structure and no formation of multinucleated giant cells, granulomas, or fibrosis [891]. Deposition of carbon powder into injured mouse lungs near the time of injury results in increased translocation of the particles to the interstitium and elicits a large increase in inflammatory cells, but does not further stimulate an ongoing fibrotic process or induce additional fibroblast growth or collagen production [851]. In one study [772], particle overload in mouse lungs produced by instilling 200 mg/kg of carbon particles caused some free carbon to cross the type I cells to reach the interstitium. These particles were later observed in peribronchial and perivascular interstitial cells [772]. In the alveoli, free macrophages were loaded with carbon but passage of these cells from airways to interstitium was never observed [772]. In another study [892], <10-micron colloidal carbon particles instilled in the lungs of rabbits shortened the transit time of alveolar macrophage-recruited neutrophils through the bone marrow from 86 hours to 71 hours. The authors concluded that the phagocytosis of colloidal carbon by alveolar macrophages releases cytokine mediators that stimulate the bone marrow to release polymorphonuclear leukocytes.
Inflammatory lung effects appear to increase when smaller particles are inhaled [4872]. In one experiment [769], ultrafine CB particles instilled intratracheally up to 0.5 mg/kg in 250-gm Wistar rats generated significant neutrophil alveolitis (alveolar inflammation) after 6 hours. The particles also produced a marked increase in lactate dehydrogenase (LDH) levels in bronchoalveolar lavage fluid and caused the greatest decrease in glutathione (GSH) in lung tissue compared to control [769]. Much larger fine CB particles similarly instilled generated no alveolitis and caused a much smaller increase in LDH and a much smaller decrease in GSH. Ordinary carbon black inhaled at 5 mg/m3 produces no significant physiological effects in rats [4859]. Fine CB shows a dose-related increasing inflammatory response. In contrast, ultrafine CB at the highest dose induces less of a neutrophil influx than at the lower dose, which the authors surmise is because particle mass dominates the response rather than surface area at higher doses. Ultrafine CB threshold dose for neutrophil influx 6 hours after instillation is 0.2 mg/kg [769]. Renwick et al [4868] found that ultrafine particles impair alveolar macrophage phagocytosis to a greater extent than fine particles compared on a mass basis.
However, at another laboratory 5 mg/kg of ultrafine and fine carbon particles were instilled intratracheally in rats and produced “little if any effect on lung permeability, epithelial marker enzymes, or inflammation, despite being given at a dose which readily translocated the epithelium and has been reported by others to cause inflammation” [857]. The authors concluded that particle surface chemistry may be more important than particle size per se, in explaining the biological reactivity of the particle [857], though a later study [5883] at this laboratory found that oxidative damage by inhaled CB particles was more strongly ameliorated by a surrogate epithelial lining fluid for coarse particles than for fine particles. In the 1990s, the relevant details of carbon surface chemistry were being investigated largely in the context of air pollution. For example, in one study [893] the inhalation of 10 mg/m3 of carbon black by mice induced no inflammatory response and had no effect on alveolar macrophage phagocytosis. But if combined with exposure to ozone at 1.5 ppm, inflammatory response was greatly enhanced and macrophage phagocytosis was significantly suppressed in comparison with exposure to ozone alone. The authors hypothesized that the carbon acts as a carrier mechanism via adsorption of ozone at the particle surface, or that O3 alters the physicochemistry of the particulate from a nontoxic to a toxic form [893]. In another study [894], a 4-hour exposure to 10 mg/m3 of carbon black aerosol at high or low humidity, assessed 3 days later, had no effect on mouse alveolar macrophage phagocytosis. But chemisorption of 10 ppm SO2 at high humidity on the carbon particles catalyzed oxidation to SO4--, a lung toxin which significantly suppressed alveolar macrophage phagocytosis [894]. There is preliminary evidence that inhaled carbon black (possibly in combination with adsorbed sulfuric acid) might promote esophageal cancer in “a handful of occupational exposures” [4817], though a competing study [2598] found no change in cell viability in lymphocytes taken from guinea pig tracheobronchial lymph nodes of animals exposed to 1500 µg/m3 diesel exhaust (carbon) particles for up to 8 weeks.
Finally, carbon-coated microbeads (Durasphere) have been tried as injectable bulking agents for treatment of urinary incontinence [4856]. But as with similar Teflon treatments (Section 15.3.4.4(2)), the particles show significant migration into local and distant lymph nodes as well as into the urethral mucosa [4856].
Last updated on 30 April 2004
{kind=link}