Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.4.2.3 Geometrical Trapping in Spleen Vasculature
The spleen has the third-highest specific blood perfusion rate in the human body, typically 48 mm3/sec-gm (~450 cm3/min) with a wide range of 10-130 mm3/sec-gm (~100-1200 cm3/min) [2727-2731]. It is probably the most likely site in the blood circulation where geometrical trapping of medical nanorobots may occur, because its microcirculation is probably the most complex of any organ in the body [5610]. The spleen serves as a sieve or filtration bed which is especially important in the clearance of rigid or less-deformable particles such as plasmodial-parasitized malarial erythrocytes [2732], red cells containing Heinz bodies [2733], poorly opsonized encapsulated bacteria [2734] (which are typically 0.6-0.8 microns in size [2735]), “sickled” cells found in sickle cell anemia patients, and colloidal particles [2736].
The spleen is a soft, purplish organ about the size of a fist whose primary role is to remove damaged, fragile or abnormal erythrocytes from the circulation. It consists of two histologically distinct areas, the white pulp (5-20%) and the red pulp (~85%) [2737-2741, 5610]. The white pulp collectively constitutes a large lymph node that performs various immunological functions such as the production of antibodies and the maturation of B- and T-lymphocytes and macrophages. Indeed, the spleen is the largest single lymphoid organ in the body – it is estimated that ~250 trillion lymphocytes/day may recirculate through the spleen of a young adult male, ~8 times more than through all lymph nodes [5616]. The red pulp acts as a blood filter to cull damaged, worn-out, or potentially dangerous cells from the blood. Most importantly, the red pulp strains out fragments of broken-up red blood cells (hemoconia or “blood dust,” probably lipid material associated with fragmented RBC stroma) and removes stiff or misshapen red cells, spherocytes (e.g., caused by IgG attachment to red cells in autoimmune hemolytic anemia [2484]), or malaria-parasitized erythrocytes [2732]. The spleen is a major site of red cell destruction in the body. It is also one of the few “dispensable” organs because mammals can survive reasonably well without one. (Because of the immune functions of the spleen, asplenic patients do have a higher risk of bacterial infections, especially from encapsulated bacteria such as Streptococcus pneumoniae, Haemophilus influenzae, and Neisseria meningitidis [5609], sometimes producing overwhelming post-splenectomy infections (OPSI) [5610] – life without a spleen is not quite as benign as life without an appendix or gallbladder.)
Blood to be filtered by the spleen enters the organ via the splenic artery. This artery ramifies into progressively smaller branches as it passes through the splenic capsule (the outer wall of the organ, comprised of collagenous connective tissue interspersed with smooth muscle) [2739]. As an arteriole enters the interior volume of the spleen, it acquires a continuous coating of lymphocytes – a thick sleeve of cells called the periarteriolar lymphocyte sheath or PALS [2742, 2743]. Arterioles continue to break into smaller and smaller vessels with thinner PALS, finally dividing into a tuft of “penicillar” arteries (so-called because of their resemblance to paintbrush bristles) [2744]. The PALS is almost wholly gone by this branching level. All of the PALS, collectively, constitute the anatomical white pulp. Upon reaching the red pulp (see below) and losing the last of the PALS, some of the penicillar arteries become sheathed capillaries whose walls are comprised of fusiform (spindle-shaped) cells. These cells are oriented parallel to the vessel axis something like a cylindrical birdcage, and are surrounded by a sheath of reticulocytes and macrophages bound together by reticular fibers [2739, 2745]. (Sheathed capillaries can in very rare cases be embolized by natural fat globules [2746]; similar embolization by a large enough population of indigestible medical nanorobots might induce clinically significant loss of function or even necrosis.) Past the sheath, these capillaries return to the normal tubular configuration and empty, along with the other capillaries, into the red pulp [2739].
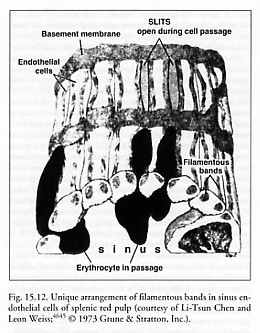
The red pulp [2737, 2738] comprises most of the splenic volume and consists of pulp cords (Billroth’s cords) and the venous sinuses (sinuses are ~30% of red pulp volume [5610]). The pulp cords make a continuous sponge-like reticular tissue, which crisscrosses between fenestrated walls of splenic venous sinuses. The cords contain erythrocytes, lymphocytes, macrophages, granulocytes, and plasma cells. The venous sinuses are 10-150 microns wide [2747] and are lined by elongated endothelial cells that resemble barrel staves ringed by hoops of reticular fibers, forming the hollow cylindrical shape of a sinus. There are tiny slits (fenestrations in the sinus wall) between these endothelial cells (Figure 15.12). The basal membranes contain actin and myosin and can probably contract to vary endothelial cell tension and thus the dimensions of the slits [5611].
The spleen filters blood via two physical pathways – “closed” and “open.” In the “closed” pathway, some capillaries terminate near the outer surface of a sinus, forcing blood to pass directly into the venous sinuses. (Arteriovenous shunts have been observed in the human spleen that are as large as 7-10 microns [2748].) In the “open” pathway, some arterioles empty blood through one or two end-pores, 3-4 microns in diameter [2749], into the pulp cords and reticular cell meshes, but not directly into the sinuses. Blood cells must then pass by open circulation through the additional barrier of the network of reticular fibers which make up the pulp cord before they can attempt to squeeze through the endothelial slits and enter the venous sinuses [2750]. Blood cells that enter a venous sinus using either pathway return to the general blood circulation, ultimately exiting the organ via the splenic vein. In humans, 90% of the blood entering the spleen takes the “closed” pathway (~2 minute transit time, directly from arteriole to venous sinuses) and only 10% goes via the “open” circulation [2751]; ~5-10% of resting cardiac output continuously passes the spleen [5612]. (The histology and microanatomy of the human spleen differs from animal spleens [5613-5615].) The slits between endothelial cells are normally closed, but widen when cells pass through them.
How large are the interendothelial slits (Figure 15.12) through which cells must pass? Fujita [2737] and Irino et al [2749] observed dumbbell-shaped erythrocytes passing through side-pores 1-2 microns in diameter, Linker [2484] gives slit width as 2 microns, and Cokelet [2752] reports that the sinus wall slits are roughly 1 micron x 6 micron rectangular openings, and about 2 microns deep.* Although red cells cannot pass through a long tube of diameter <~2.3 microns without plugging the tube (Section 8.2.1.2), careful experiments have revealed that normal human erythrocytes can traverse a rigid micropore with a diameter as small as 1 micron if the pore depth is very shallow (e.g., 0.4 micron [2753]) with a driving pressure of only ~1.5 mmHg differential [2754], or even as small as 0.5 micron during erythrocyte diapedesis [2755]. Transit through the slit is slow, ~10 sec or longer for healthy red cells [2752], somewhat slower in rat [2756]. The mean residence time of a human erythrocyte in the red pulp is ~66 sec with an average velocity of ~0.25 micron/sec [2752]. Because the percolation of blood through the reticular meshwork between capillary endings and venous sinuses is so slow, the red pulp concentrates blood to twice the normal arterial hematocrit, e.g., an intrasplenic hematocrit of ~78% [2757].
* An early oft-cited electron microscopy study by Chen and Weiss [4645] reporting an interendothelial slit size of 0.2-0.5 micron in width, 2-3 microns in length, and 3-5 microns in thickness was for phenylhydrazine-poisoned Sprague-Hawley albino rat spleen, not healthy unpoisoned human spleen. Human erythrocytes have a much larger mean cell volume (MCV) of 94 micron3 (Section 8.2.1.2) than the reported MCV of 59.7 micron3 for rat erythrocytes [4646], and phenylhydrazine can dramatically affect cell volume [4646] and thus, quite likely, measured absolute slit width as well.
In the red pulp, deformed or injured red cells cannot navigate the splenic sinusoids successfully, leading to sequestration and phagocytosis (Section 15.4.3.2.4). For instance, red cells containing Heinz bodies (granules due to hemoglobin damage) have difficulty traversing even shallow pores ~3 microns in diameter [2733], and relatively rigid spherical Staphylococcus aureus bacteria of diameter 2-3 microns have been found trapped in mouse spleen [2758]. In this manner, the spleen monitors for abnormal cells in blood by geometrical sieving in venous sinuses and by surveillance of surface antigens by mononuclear phagocytes. Experiments show that polystyrene microspheres 5 microns in diameter injected into rat splenic artery are trapped mainly in the red pulp [2759]. One possible additional minor complication is the observation that rat and mouse spleen capillaries apparently can experience spontaneous cyclic contractions of capillary walls to as narrow as 1 micron in diameter [2760]. Cycles average 1 minute in length (range 12-180 sec), with closure occurring in 2-12 sec during the cycle, lasting for <1-60 sec. However, the author is unaware of any reports of such contractions in human spleen.
Can medical nanorobots pass through the venous sinus slits? These slits are optimally designed to trap rigid particles. For instance, Heinz body fragmentation occurs when rigid particles of oxidized hemoglobin are torn from affected red cells as they circulate through the spleen [2761]. Thus the simplest device design rule would be that non-organic nanorobots should possess at least one physical dimension of ~1 micron or less, or should employ mechanical assistance [2762] or a metamorphic surface (Section 5.3) capable of deforming the entire device to a width of ~1 micron or less in at least one dimension during slit passage.
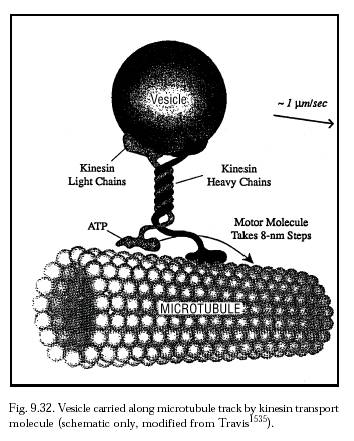
It may be useful for bloodborne nanorobots larger than ~1 micron in any dimension to incorporate explicit splenic passage protocols and splenofenestral motility mechanisms in their design [2762]. The situation that such a nanorobot may face in traversing the spleen might best be understood by analogy to the recorded journey of a typical healthy erythrocyte through the red pulp. In one experiment, an analysis of three 70-micron RBC paths through open red pulp showed characteristic stop-and-go motion, with RBCs observed spending 0.015-9.71 sec in any 7-micron segment, despite steady perfusate velocity [2757]. At some locations RBCs adhered to reticular cells or fibers by point attachment, and at others they became caught over fibers. But in general RBCs were detained in the reticular meshwork more by surface interactions rather than by narrow channel restrictions [2757]. Appropriate splenofenestral passage techniques for medical nanorobots might range from relatively simple motile semaphoric surface arrays (Section 9.4.5.3) over individual step lengths perhaps equivalent to the 8-nm kinesin motor molecule steps (Figure 9.32), to more complex brachiation mechanisms [2762] (Section 9.4.4.2). Lymphatic-return pathways might also be feasible from the spleen [2743]. A more aggressive solution would be to deploy in advance a small number of specialty nanorobots to selectively block the entrance to the splenic artery during the nanomedical procedure, physically denying entry to therapeutic nanorobots while simultaneously allowing other blood components to pass normally. One such nanorobotic “vascular gate” (see Chapter 19 for details) installed across a 6-mm diameter artery could be established using a sheet of ~107 nanorobots each having a (~2 micron)2 patrol area. The gate would allow non-nanorobotic particulate matter to pass, selectively filtering out only nanorobots; if the vascular gate aggregate consists of vasculomobile nanorobots, then at mission’s end the aggregate can disassemble itself and “walk away” without creating a large nanorobotic embolus that could infarct the spleen. (Vascular gates seem workable for nanorobots on short-term missions but might not be appropriate for long-term missions where nanorobots are performing surveillance functions such as early disease detection.) This avoids the even more radical (though survivable) conventional options of splenic embolization [2763] or splenectomy followed by autologous splenic reimplantation.
Some blockage of the reticular meshwork by insoluble particles is tolerable and presents only minor health risks. For example, experiments with 3-micron latex spheres in rats found these particles to be safe imaging agents or drug delivery systems for spleen or liver [2679], even though 3.4-micron microspheres pass readily through the lungs and are trapped in the spleen with bloodstream half-lives of 1.62 minutes and 1.72 minutes in venous and arterial circulation, respectively [2686]. Acute hemodynamic toxicity as a result of vascular occlusion is a function of total microsphere volume administered, with an inverse relationship between sphere diameter and hemodynamic toxicity [2679]. Additionally, there is some evidence that up to 90% of the blood flowing into the spleen may pass through a region bordering the white pulp known as the perimarginal cavernous sinus plexus (comprised of large flattened open spaces up to 300 microns x 1000 microns in area and 30-100 microns thick), bypassing the narrow-gauge filtration beds of the red pulp [2745].
Last updated on 30 April 2004
{kind=link}
{kind=link}