Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.5.7.2.3 Resident Transmembrane Penetrators
In some applications it will be desirable to establish a plasma membrane penetration for the duration of the nanomedical mission. Such applications may involve rods, cables or tubing that must pass from the extracellular to the cytoplasmic spaces through a fixed sheath, or artificial diamondoid sensors, pumps, or other mechanisms that must be installed in the plasma membrane wall and perform some useful function throughout the mission (e.g., external chemical sensors during cytocarriage; Section 9.4.7.5) without eliciting unwanted mechanosensitive biological responses.
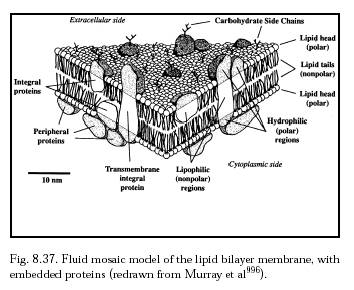
The existence of natural transmembrane integral proteins (Figure 8.37) and artificial transmembrane ion channels [5426-5436], synthetic pores and porins [5437-5439], engineered antibiotic-based channel-forming peptides [5440-5443], and artificial organic nanotubes [692, 5444] implies that it should be possible to design, install and stabilize artificial transmembrane penetrator nanodevices. Such devices may anchor themselves in the membrane using an amphipathic coating similar to that employed by integral proteins – e.g., a hydrophilic (polar) coating above and below the plasma membrane, corresponding to the position of the polar lipid heads in the membrane, and a lipophilic (nonpolar) coating for the midmembrane region, corresponding to the position of the nonpolar lipid tails in the membrane. This gives the surface of each nanodevice the maximum membrane affinity when the device is properly positioned, and anchors it in place. Deployment of these anchoring coatings may be reversibly controlled by a nanorobot using an array of presentation semaphores (Sections 5.3.6 and 9.4.5.3). The hollow interior of a penetrating sheath should provide a low-friction interface with a fluid-tight seal (Section 10.3.4; and also Section 11.4.2 in Drexler [10]) so that forces from a cable, rod, or other object sliding through the sheath are not efficiently transmitted to the sheath and thence to the membrane and its attached cytoskeletal components. Hamill [personal communication, 2002] suggests that “nanorobots might be able to insert themselves into selective areas of the plasma membrane (e.g., of polarized cells) by hitchhiking in the trans-Golgi vesicles that are used to traffic/direct newly synthesized membrane proteins to specific regions of the cell surface.”
Hamill and Martinac [5640, 5641] have found that sub-nanometer changes in bilayer thickness can switch gramicidin A from a stretch-activated to stretch-inactivated channel. As they note, the presence of “a bilayer-controlled switch in signaling by a mechanotransducer channel emphasizes that the bilayer is much more than a neutral solvent [and] may actively modulate the specificity and fidelity of signaling by membrane proteins. This feature, in combination with protein-related factors (e.g., oligomerization state and cytoskeleton association) that determine not only protein recruitment into lipid microdomains but also the dynamic organization of the bilayer itself, indicates a dynamic reciprocity in lipid-protein interactions that is presumably necessary for the higher-order spatial and temporal control of signaling.” The presence or movements of, or the forces applied by, a resident transmembrane penetrator could alter the thickness of adjacent bilayer, locally modifying the behavior of nearby mechanosensitive or mechanotransducing integral proteins. These modifications could significantly impact cell function – possibly including cell responses accompanying membrane resealing [5640] – if substantial numbers of artificial penetrators are resident (especially if closely grouped) on the plasma membrane of a single cell. Switch effects presumably may be minimized by avoiding both static and dynamic hydrophobic mismatch [5640, 5641] between the hydrocarbon portion of the lipid bilayer (which may vary for different cell types) and the hydrophobic exterior surface of the penetrator.
Other complicating factors include the constant turnover of the plasma membrane (Section 8.5.3.2) and the likelihood that a cytoskeletally-unanchored penetrator might be dragged along with the usual lateral motions of membrane raft microdomains [4247, 4248] and untethered integral proteins [4249] such as red cell aquaporins [4250]. Nevertheless these motions should be confined to submicron domains on the surfaces of many cells [4251-4255]. Assuming the nanodevice penetrators are not very numerous in the cell wall, they should not substantially alter plasma membrane fluidity or viscosity especially if differential shear forces are low, e.g., <1 N/m2 [4256].
Last updated on 30 April 2004
{kind=link}