Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.4.3.1 Phagocytes, Phagocytosis, and the RES
There are two broad classes of “professional” phagocytes. First, there are the granulocytes or polymorphonuclear leukocytes (“PMNs” or “polys”), having horseshoe-shaped multi-lobed nuclei, that circulate in the blood. The PMNs include the neutrophils (the most common granulocyte), basophils, and eosinophils. Second, there are the agranulocytes or mononuclear phagocytes (“MNPs”), which have only one nucleus. The mononuclear phagocytes include the monocytes (in the bloodstream), lymphocytes (mostly resident in lymphatic tissues [5670]), and macrophages (monocytes that have left the bloodstream, transformed, and settled in the tissues). Many other cells in the body, such as endothelial cells [2833-2836], fibroblasts [778], osteoclasts [2837], pericytes [2833, 2834], and platelets [775, 868, 875, 881-883, 885] have some phagocytic activity, and even thyroid and bladder epithelial cells phagocytize erythrocytes in vivo [2867]. But only the neutrophils and macrophages are good enough particle scavengers to be considered professional phagocytes [2838].
Phagocytic cells are normally inactive until an apoptotic ([5765-5767, 6065]; Section 10.4.1.1) cell* or a foreign cell or particle is encountered, which activates them [234, 647]. Activation is characterized by a change in metabolic activity (e.g., an increase in oxidative metabolism or “respiratory burst”**) and in cell shape. When contacting and recognizing a foreign particle through antigen (opsonin)-membrane receptor binding, the phagocyte plasma membrane develops a dimple (invagination). The particle is drawn inside and the dimple closes, often pinching off to form a small vacuole or phagosome. This traps the particle inside the cell, surrounded by everted cell wall membrane. The phagosome then forms a phagolysosome by merging with a lysosome, whose contents (including degradative lysozymes) are released into the smaller vacuole, attacking the enclosed foreign or denatured proteins [234]. There are ~200 cytoplasmic lysozyme-containing granules per neutrophil [2839]. The ensuing oxidative burst [2875, 2877] produces hydrogen peroxide and superoxide anions [2840] which act nonspecifically to digest nonproteinaceous foreign materials that cannot be digested by lysozymes.*** Afterwards the phagolysosomal vacuole may be absorbed or released to the outside at the cell’s outer surface (exocytosis; Section 8.5.3.7), producing a large membrane flow. For example, in cultured macrophages an amount of membrane equal to the entire surface area of the cell is replaced in ~1800 sec [2841], and macrophages may ingest up to ~25% of their volume per hour [526]. For macrophages ranging from 25-30 microns in diameter [2844], these numbers imply a maximum particle-wrapping phagosomal membrane recycling rate of 1.1-4.4 micron2/sec and a maximum volumetric ingestion rate of 0.6-4.6 micron3/sec, which in turn suggests a theoretical maximum particle phagocytosis rate of one ~1-micron3 particle per second, per macrophage.
* Interestingly, phagocytes can also induce apoptosis (cell death) in seemingly healthy cells [5262] (or at least in cells whose unhealthy state is too subtle for us to detect today).
** Respiratory burst refers to an increase in oxidative metabolism, including oxygen consumption, that occurs after the phagocyte ingests the opsonized particle. The major products of respiratory burst in cells are superoxide anion, singlet oxygen, hydrogen peroxide, and hydroxyl free radical [5617].
*** However, at least one recent study [4629] suggests that activated proteases could be mainly responsible for bacterial destruction inside neutrophils, with toxic reactive oxygen species and myeloperoxidase-catalyzed halogenation (iodination) playing lesser roles.
Of course, the normal rate of phagocytosis is low in relation to this theoretical maximum capacity [2845]. If the chemical composition of foreign particles allows their degradation, they are destroyed. But if the particles are resistant to digestion, they are retained within the phagocyte in isolated phagosomes [2842] and thus are effectively removed from further interaction with the host [2843]. Typically:
(1) internalized albumen is digested inside macrophages with a half-life of 13-23 minutes [2846];
(2) rat macrophage digestion of opsonized red cells and ghost red cells is complete in 24 hours and 3 hours, respectively [2847];
(3) Kupffer cells digest organelle membrane proteins with a half-life of 1.5-2 hours and lipid components with a half-life of 2.0-3.5 hours [2848];
(4) surface-bound immune complexes are digested by macrophages with a 15.5-hour half-life at 37 oC [2849];
(5) the digestive half-life for mitochondria by rat liver Kupffer cells is 3-4 hours, and 8 hours for microsomes [3664];
(6) tooth amoebas digest human erythrocytes with a 2-hour half-life [2850]; and
(7) ribosomes require >24 hours to be fully digested inside rat Kupffer cells [2851].
Neutrophils, typically 8-10 microns in diameter, are specialized to find and phagocytize bacteria. They can also detect and become activated by foreign particles such as splinters and wear debris. The process of inorganic particle detection and PMN activation remains incompletely understood, but appears to be guided by several mechanisms including particle chemical composition (chemotaxis); local pH differences; electrochemical factors associated with the particle and its surroundings; opsonization (e.g., by complement; Section 15.2.3.2) of the particle [234]; and most importantly by the availability of excess plasma membrane area [2852]. However, medical nanorobots can probably be engineered to avoid PMN activation. This is because micron-size diamond particles do not activate complement [1642], do not produce neutrophil chemotactic activity [222, 633, 639], and do not stimulate lysozyme degranulation in neutrophils [640]. Diamond particles are ingested by PMN cells (Section 15.3.1.4) – admittedly, an event that could be significant enough to prevent a diamondoid nanorobot from completing its mission – but there is no significant further biological response after such ingestion. In one experiment [640], 4- to 8-micron diamond crystals present at up to ~0.2% Nct (nanocrit, % by volume; Section 9.4.1.4) in culture were ingested by PMNs without negative effect. In another experiment [633], 3-micron diamond crystals at 2 mg/cm3 (~0.06% Nct in culture) were phagocytized by 21% of the neutrophils present at 7250 cells/mm3 after 45 minutes, but again no chemotactic activity was generated. Eosinophils are similar in structure and function to the neutrophils, and can also phagocytize antigen-antibody complexes [234], but no antibodies have yet been raised to diamond (Section 15.2.3.3). Neutrophils and eosinophils are the first active line of defense against foreign material in tissue – an “emergency squad” whose duties are later supplanted by monocytes [234]. Leukocytes are end-state cells that cannot replicate by division (mitosis). They have a lifespan of a few hours in blood or few days in tissue, and the cells die rapidly after fulfilling their phagocytic function [234]. Heat production rises from 9 pW/cell in unstimulated human neutrophils up to 28 pW/cell during phagocytosis, with the rise proportional to the number of particles ingested [2853].
The monocyte is the largest freely-circulating leukocyte, up to 12-15 microns in diameter [2844], which transforms into a macrophage upon its permanent migration into tissue. There is also a pool of transformable monocytes already present in the tissues [234]. The estimated basal whole-body monocyte production rate is ~11,000 cells/sec for the average adult human [2854]. Monocytes circulate in the bloodstream for 8-70 hours [2854]. During this time they enlarge, migrate into the tissues, and after 8-12 hours differentiate into specific tissue macrophages [2855]. MNPs can actively phagocytize and digest foreign materials [2856], and can also synthesize and release a wide range of biochemical factors to mediate the local activities of other cells such as lymphocytes and fibroblasts [234]. Macrophages have a maximum lifespan of several months [2004], replacing themselves at the rate of ~1%/day [2855]. They typically measure 25-50 microns in diameter [2844]. Some macrophages can multiply by mitosis, or can fuse to create the multinuclear foreign body giant cell (FBGC) as a direct response to larger foreign particles [2857, 2858]. FBGCs can reach up to 80 microns in diameter and are found primarily in foreign body (Section 15.4.3.5) or implant sites [234].
The mononuclear phagocyte system (MPS) [2859] – classically known as the reticuloendothelial system (RES) [2860] – is the primary active system in the human body responsible for the removal of old and damaged cells, cellular debris, pathogens and other foreign particles from the circulation [2861]. The RES is composed mostly of fixed, but some wandering, cells, derived from the original bone marrow monocyte [2862]. RES phagocytes are found in large quantities in the spleen (sinusoidal cells), lymph nodes (lymphocytes), and lungs (dust cells or alveolar macrophages). But the Kupffer cells in the liver represent about 50% of all macrophages in the human body. Other macrophages are present in smaller numbers in the blood (monocytes), brain (microglia), kidney (mesangial cells), bone marrow, adrenals, thymus, mucous membranes, serous cavities, breast, placenta, and connective tissue (histiocytes). The human RES consists of at least 200 x 109 phagocytic cells [2863].
The presence and activity of phagocytes is particularly related to the presence of small particles. For example, 0.325-micron PMMA particles stimulate cytokine release in vitro by human macrophages at concentrations exceeding 1010-1011 particles/cm3 [2864]. Maximum stimulus occurs at average particle sizes in the 0.1-2.0 micron range [234, 2865-2868], though phagocytosis is often said primarily to involve the uptake of particles >0.5 micron in size [2867].
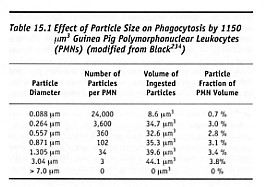
At the larger sizes, murine bone-marrow macrophages which are 13.8 microns in diameter can ingest IgG-opsonized beads >20 microns in diameter [2876]. Ingestion of an opsonized 15-micron particle requires 30 minutes to reach completion [2876], whereas ingestion of a lymphocyte by a macrophage was observed to require only 3 minutes (with dramatic shape changes, including formation of a pseudopod 155 microns in length) [2869]. Maximum neutrophil pseudopod extension speed is ~50 microns/minute [2870]. Another experiment [784] with cultured murine macrophages found that inert carbon fiber-reinforced carbon particles up to 20 microns in diameter are phagocytosed. Larger particles are not phagocytosed but become surrounded by aggregations of macrophages, some of which migrate onto the particle surfaces [784]. Presented with these larger particles, several macrophages can fuse to form a giant cell, which subsequently may be cleared into the lungs via the lymphatics and expectorated or passed into the intestines for elimination [2871]. Still larger particles (>50 microns) generally don’t excite a reaction greater than bulk materials [234] unless they possess a dimension in the smaller size range – e.g., long slender fibers [2878]. The largest particles that neutrophils can ingest are somewhat smaller than those for macrophages. For example, Table 15.1 shows that guinea pig PMNs cannot absorb particles >~7 microns in diameter. Human blood monocytes readily ingest inert 0.39-micron particles, rarely ingest 1.52-micron particles, and never ingest 5.1-micron particles [2868]. Individual macrophages rarely ingest particles larger than ~5 microns [1074, 2864]: such particles are ingested by FBGCs (foreign body giant cells). Ingestion may proceed differently in larger cells. For instance, in amoebas, larger particles are taken up singly while smaller particles are accumulated outside the cell. When a critical volume is reached, the small-particle aggregate is absorbed all at once [234, 2872, 2873].
A phagocytic cell can become activated by a failure to digest indigestible or toxic particles (e.g., silica crystals absorbed by alveolar macrophages [175]). This results in the external release of lysozymes and oxidative products, a process called frustrated phagocytosis [2874-2879]. When this occurs, the offending particle is often expelled unchanged from the dying macrophage, followed by ingestion by another macrophage which itself will be killed. This process produces a continuous inflammation with masses of dying and dead neutrophils or macrophages, as well as a cellular debris accumulation (caseation) or “pus” resembling that which accompanies massive bacterial infection [234]. Fortunately, as with neutrophils, 2- to 4-micron diamond particles can be ingested in large quantities by macrophages without harmful effect on the cells [652]. Phosphatase enzyme discharged into diamond-containing phagosomes does not escape into the macrophage cytoplasm or nucleus [652]. In another study, 2- to 15-micron diamond particles in serum-free suspension at 0.5 mg/cm3 (~0.01% Nct in culture) induced no change in monocyte morphology, indicating no phagocytic activation by the diamond [641].
Mean rates of phagocytosis up to 2.5 particles per hour were observed for cultured murine macrophages ingesting 8- to 20-micron inert carbon particles [784]. A second experiment [1074] found only 10% mortality among murine macrophages that had ingested up to 2500 0.6-micron alumina ceramic particles (~10% of cell volume), but >30% mortality for particles >2 microns in diameter at high concentrations.* Aggregated serum albumin is cleared at rates similar to rates for metal, dye, and other inert particles. The whole-body maximum clearance rate of aggregated albumin by the human RES has been determined to be ~1.07 mg/kg-min, or ~1.25 mg/sec for a 70-kg man [2863].
* Interestingly in macrophages, as in PMNs, maximum phagocytic capacity appears to be limited by the amount of available membrane rather than by the number of surface receptors [2876].
In the natural course, inert particles, once ingested, are retained in isolated phagosomes for the life of the phagocyte. Phagocytes that have ingested too many inert particles lose their phagocytic function, a process called blockade [872, 873, 1391, 3631-3634] (Section 15.4.3.6.10), and shorten their lifespan. Thus massive tissue overloads of chemically inert nanorobots might blockade or even kill large numbers of phagocytic cells if the internal accumulation of foreign matter volume becomes too great (perhaps ~10-20% of total phagocyte volume; Section 15.6.3). In the experiment using carbon particles [784], cells presented with a large excess of inert particles became rounded and detached from the substrate, and some cells underwent lysis. Inert particles released into the intracellular space due to the lysis of a blockaded phagocyte will either be re-ingested by another phagocyte, or will be swept into the lymphatic filtrate and eventually sequestered in lymph nodes (Section 15.4.3.4) or granulomatized (Section 15.4.3.5).
As a general rule, phagocytic response is decreased with increasing particle size [2880], producing differences in histological reaction [644] and cytokine production [645]. For instance, particles of hydroxyapatite larger than 15 microns are not taken up intracellularly by monocytes [641] and thus appear relatively inert [2881]. But <15-micron particles of similar composition, when added to serum-supplemented cultures, stimulate monocytes to produce bone resorptive cytokines [2882, 2883], which has been corroborated under serum-free conditions [2884].
The influence of surface charge on phagocytosis is less clear. One study found no significant difference between anionic and cationic surfaces [2865]. Other experiments with polystyrene microspheres having macromolecule-modified surfaces produced different clearance and organ deposition patterns for negatively or positively charged particles [2688], and suggested that positive charges increase phagocytic uptake while negative charges reduce uptake [2880]. Positively-charged particles tend to accumulate in the lungs, whereas negatively-charged particles tend to accumulate in the liver, with very few found in the lungs or spleen [2688]. But total RES distribution is not changed, relative to non-RES tissues, despite these differences in organ distribution, so the end result of surface charge alteration is mainly a redistribution within the RES [2885].
Hydrophilic particles are phagocytosed less [2880]. For instance, microspheres with hydrophobic surfaces are more readily phagocytosed than those with hydrophilic surface in murine peritoneal macrophages [2865]. Surface hydrophobicity appears to be a critical determinant in the opsonization process and in the subsequent uptake of particles by the RES [2886]. Particles with hydrophilic surfaces are rendered more hydrophobic by the adsorption of IgG and can then become phagocytized by macrophages [2874]. In contrast, hydrophobic particles are taken up by macrophages without the need for opsonization [2764]. In one experiment, the least phagocytosis was observed for cellulose microspheres with non-ionic hydrophilic surfaces [2865]. Since all these properties are readily controlled by design, medical nanorobots probably can be given the ability to evade, or if necessary, to escape from (Section 15.4.3.6), the RES.
Particles smaller than 0.5 microns in diameter that are hydrophobic – such as aggregated LDL, microcrystalline cholesterol, polystyrene microspheres, or hydrophobic gold – may trigger “patocytosis.” Patocytosis is a unique macrophage endocytosis pathway in which external particles induce and enter a labyrinth of internal membrane-bound compartments that remain connected to the phagocytic cell surface [2887]. Hydrophobic polystyrene microspheres larger than 0.5 microns enter macrophages in the usual manner, via phagocytosis [2887].
Interestingly, tissue cells contain endogenous adjuvants in their cytoplasm that when released (e.g., due to cell injury or death by apoptosis) markedly augment the generation of CD8 cytotoxic T lymphocyte responses to particulate and cell-associated antigens but not to the same antigens in soluble form [5047]. This is a different mode of action than a classical immunostimulant or bacterial adjuvant such as Freund’s. Experimental co-injection of cytosol and fluorescent particles increases the accumulation in the draining lymph node of dendritic cells and macrophages that contain phagocytosed particles and that express high levels of costimulatory molecules [5047]. As a result, attempts by phagocytes to trap immune-visible medical nanorobots may become more urgent in the immediate locale of cell trauma or apoptosis, unless the endogenous adjuvant molecules are extracted or metabolized by the nanorobots.
Last updated on 30 April 2004
{kind=link}