Nanomedicine, Volume IIA: Biocompatibility
© 2003 Robert A. Freitas Jr. All Rights Reserved.
Robert A. Freitas Jr., Nanomedicine, Volume IIA: Biocompatibility, Landes Bioscience, Georgetown, TX, 2003
15.5.7.1 Mechanical Interactions with Glycocalyx
Many disease processes are known that involve damage to the glycocalyx [4197-4208], including some bacteria that phagocytose [4209] or otherwise destroy [4210-4213] the host cellular glycocalyx during an infection. Damage to the glycocalyx creates conditions that favor the binding of immune complexes, complement activation, and intravascular coagulation, with loss of gradients between blood and parenchyma [4214]. Desialylated glycocalyx of endothelium also allows an increased rate of endothelial cell detachment from arterial walls [4215].
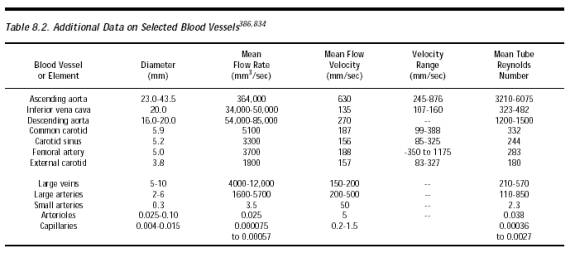
Nanorobots that rely upon absorption of local oxygen and glucose for their power supply (Section 6.3.4) or whose missions include extensive small-molecule exchanges with the environment (Chapter 19) may have ~104-105 molecular sorting rotors (Section 3.4.2) embedded in their exterior surfaces [2762, 3573]. These spinning sorting rotors are unlikely to cause direct physical damage to formed blood elements for several reasons. First, rotors are atomically smooth and recessed into the housing, reducing physical contact with colliding surfaces and eliminating potential nucleation sites that may trigger blood clotting, gas embolus formation, or foaming. Second, only a small fraction of all available sorting rotors may be actively spinning at one time, further reducing the likelihood of physical trauma. Third, such limited physical contact, when it occurs, should be relatively benign. Maximum rotor rim velocity of 2.6 mm/sec is less than 1% of mean aortic blood velocity and lies only slightly above maximum capillary flow speed (Table 8.2).
Could the glycocalyx strands that are present at all tissue and nontissue cells surfaces get trimmed, even by a recessed sorting rotor? Nanorobot sorting rotor binding sites for small molecules (<20 atoms) involve pockets measuring <2.7 nm in diameter ([10], Section 13.2.1.a). These pockets are too small to physically accommodate the 10-20 nm thick plasma membrane or the main body of the glycocalyx projections that typically measure 5-8 nm thick and 100-200 nm long [4216], and consist of glycoproteins comprised of 10,000 atoms or more. While an occasional sugar residue may get clipped, binding sites can be designed for maximum steric incompatibility with glycocalyx glycoproteins and proteoglycans, further minimizing the opportunities for trimming. Note that clipping a covalent C-C, C-O, or C-N bond probably requires a clipping energy >500 zJ/molecule ([10], Table 3.8), but sorting rotors designed to pump against pressures of ~30,000 atm can only apply ~100 zJ/molecule (i.e., per binding site). An accidentally-bound glycocalyx moiety seems more likely to jam the rotor than to be clipped off by the rotor. If this happens, the result may be a glycocalyx-tethered nanorobot, in which case a rotor-dejamming protocol* is needed to free the trapped nanorobot.
* One obvious backflushing procedure would use follower rods to affirmatively push unwanted ligands out of the binding pocket. Additionally, some molecular sorting rotor designs ([10], Section 13.2.1.d) assume a compliant mechanical coupling that permits the rotor to spin backward a short distance as if in free rotational diffusion, thus allowing improperly bound ligands to be freed.
But what if glycoproteins can be clipped? Consider a population of Nbot spherical nanorobots of radius Rbot, the surface of each is covered with a fraction frotor of sorting rotors each of face area Arotor with binding site aperture area Abind, which are resident at nanocrit Nct in a blood compartment of volume Vblood in which each nanorobot experiences cbot collisions/sec with glycocalytic biological surfaces of total area Aglyx, where Nbot = 3 Nct Vblood / (4 pi Rbot3) and the fraction of rotor surface that is binding site aperture is krotor = nrotor Abind / Arotor, for rotors with nrotor binding sites always exposed. The rate at which biological glycocalyx is encountered by the collective binding site aperture area of the entire fleet is Sbot ~ pi Rbot2 cbot Nbot krotor frotor (m2/sec), so each glycoprotein strand is presented to a nanorobot binding site aperture once every Aglyx/Sbot seconds. If we assume that:
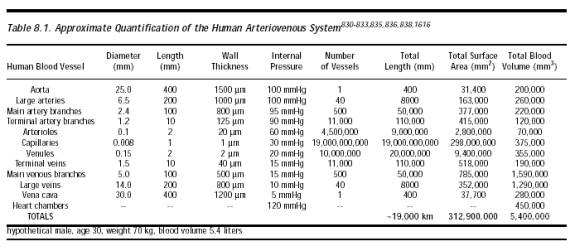
(a) one in Nenc such encounters positions a glycoprotein strand moiety such that binding with it would be geometrically possible; and
(b) epsilonrotor is the relative affinity of the binding site for glycoprotein moieties (e.g., glucosyl, sialyl) properly presented to it as compared to desired targets such as O2 molecules – that is, the rotor’s specificity for glycocalyx – and
(c) treplace is the natural replacement time for damaged glycoprotein strands in the glycocalyx;
then epsilonrotor <~ kappax Aglyx Nenc / (treplace Sbot), where kappax is the maximum permissible fraction of damaged glycocalyx during the time treplace. For this analysis, we take Vblood = 5400 cm3, Rbot = 1 micron, frotor = 1, cbot ~ 100 collisions/sec (Section 15.5.5.2.1), Arotor = 98 nm2 (Section 3.4.2), Abind = 0.033 nm2 for an oxygen molecule (the most common molecule likely to be sought by medical nanorobot exterior rotors), Nenc ~ 10, and Aglyx ~ 4410 m2 (which is the sum of ~3850 m2 for 28.5 trillion RBCs (Section 8.5.1), ~310 m2 for the vascular endothelium (Table 8.1), ~210 m2 for 0.7 trillion WBCs (Section 8.5.1), and ~40 m2 for 2.1 trillion platelets (Section 8.5.1)). Since red cells are the most commonly impacted and do not self-repair, the worst-case replacement time is the mean RBC lifetime of ~120 days, or treplace ~ 107 sec. Natural rates of glycocalyx damage are just starting to be precisely quantified [4217, 4218], so for the present analysis we assume that kappax ~ 0.01 (1%) is an acceptable glycocalyx damage rate.
At Nct ~ 0.10 (10%), the largest plausible nanocrit (Section 9.4.2.6), the required relative rotor affinity for glycoprotein moieties vs. oxygen molecules is epsilonrotor <~ 10-6. Drexler [10] notes that “analogies with antibodies suggest that an inward-transport rotor can deliver impurity fractions of 10-4 to 10-9, depending on affinities, specificities, and the concentrations of the effectively competing ligands.” Hence, it appears likely that rotor systems can be designed to achieve acceptable glycocalyx damage rates of kappax <~ 1% – if clipping damage is even possible at all. This conclusion is further reinforced when we consider: (1) that a more typical diagnostic or therapeutic medical nanorobot dose will be ~1 terabot (Nct ~ 0.01%), not ~1000 terabots (Nct ~10%) as assumed above; (2) that a more typical mission time may be only hours in duration (~104 sec), rather than the RBC lifetime (~107 sec) as assumed above, and may be comparable to or less than the time in which many tissue cells replace their glycocalyx or are retired, e.g., 103-106 sec*; and (3) that recessed access ports or protective cowlings near the binding sites might increase Nenc to 100 or better. All of these factors combine potentially to reduce the aforementioned worst-case damage rate by up to 7 orders of magnitude.
* Schistosome parasites can shed some tegument-bound complexes in only ~1200 sec [503] to 3600 sec [4219]. Plasma membrane turnover rate is ~1800 sec for macrophage [2841] and ~5400 sec for fibroblast [526]. Cholesterol turnover rate in RBC membrane is ~7200 sec [4220]. Membrane phospholipid half-life averages ~10,000 sec [353]. Neutrophil lifespan in blood is ~11,000 sec [234]. Enterocyte glycocalyx is renewed in 14,000-22,000 sec, as vesicles with adhered bacteria are expelled into the lumen of small and large intestine [4221]. Some schistosome membrane antigen turnover may require from 68,000 [4222] to 160,000-430,000 sec [493]. Typical protein turnover half-life is ~200,000 sec [353, 4223]. Cell turnover time is ~86,000 sec in gastric body, ~200,000 sec for duodenal epithelium, ~240,000 sec for ileal epithelium, and ~400,000 sec for gastric fundus [1841]. Neutrophil lifespan in tissue is ~260,000 sec [234]. Glycocalyx turnover in rat uterine epithelial cells is ~430,000 sec [4224]. Platelet lifespan is ~860,000 sec [4095].
The glycocalyx cell coat is a secretion product incorporated into the plasma membrane that undergoes continuous renewal. Thus any trimmed glycocalyx glycoproteins from tissue cells would be rapidly replaced via biosynthesis in the ribosomes of the endoplasmic reticulum, followed by final assembly with the oligosaccharide moiety in the Golgi complex and subsequent export to the plasma membrane [4225]. Glycoprotein strands or stray sugar residues released into the extracellular medium as a consequence of such trimming are nonimmunogenic and would be quickly metabolized. However, it is possible that nearby parasites could absorb this released material onto their surface, affording themselves some camouflage protection (Sections 15.2.3.4 and 15.2.3.6) against host immune defenses [4226].
Last updated on 30 April 2004
{kind=link}
{kind=link}